
artikel
De reeds verschenen artikelen uit deze serie zijn gepubliceerd op bl. 1626, 1627, 1725, 1835, 1951, 2064, 2164, 2262, 2365 en 2469 van de vorige jaargang.
Een cel kan de functie van een andere cel beïnvloeden door signalen uit te zenden die door de ontvangende cel ‘begrepen’ kunnen worden: wij spreken dan van prikkeloverdracht of signaaltransductie. Deze informatieve signalen kunnen zowel elektrisch als chemisch van aard zijn. In dit hoofdstuk zal de nadruk op chemische prikkeloverdracht liggen. Chemische signalen kunnen zowel lokaal (paracrien) als op afstand, na transport via de circulatie (endocrien), actief zijn. Overdracht van chemische signalen van de ene cel naar de andere vindt plaats door binding van signaalmoleculen (liganden) aan specifieke receptormoleculen. Door binding van de ligand wordt de receptor geactiveerd, waarna signaaloverdracht kan plaatshebben. Polaire liganden, zoals peptidehormonen, kunnen de celmembraan niet passeren en werken via binding aan receptoren in de celmembraan. Deze binding leidt tot intracellulaire veranderingen in de concentratie van ‘second messengers'cdie de ’boodschap‘ overbrengen. Apolaire liganden, zoals steroïde hormonen, kunnen de Celmembraan wel passeren en werken via binding aan intracellulaire receptoren. De eiwitten die na activering via één van de routes van prikkeloverdracht de cellulaire reactie op een prikkel teweegbrengen, worden effectoren genoemd.
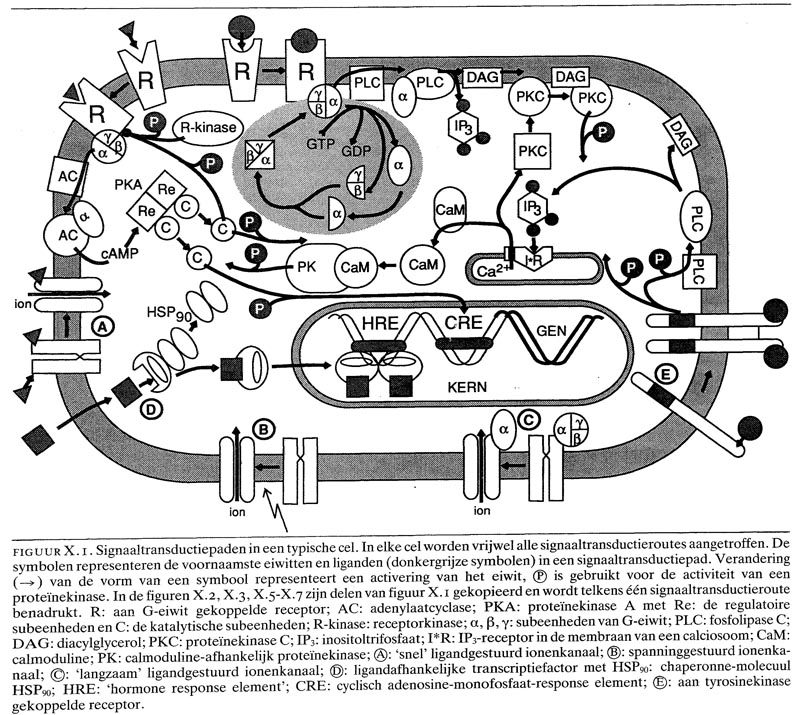
X. 1. routes van signaaltransductie
Het aantal verschillende mechanismen waardoor signaaloverdracht kan plaatsvinden, is betrekkelijk beperkt. De specificiteit van de cellulaire reactie op een extracellulair signaal is binnen elk der routes meestal te herleiden tot verschillen in het ligand-bindende domein van de receptormoleculen. Tot nu toe zijn zes verschillende typen receptoren geïdentificeerd (figuur X.1):
– ligandafhankelijke transcriptiefactoren
– aan G-eiwit gekoppelde receptoren, namelijk
– van cyclisch adenosine-monofosfaat (cAMP) of cyclisch guanosine-monofosfaat (cGMP) afhankelijke proteïnekinasen
– fosfolipide-afhankelijke proteïnekinasen
– Ca2calmoduline-afhankelijke proteïnekinasen
– ‘langzame’ ligandgestuurde ionenkanalen
– spanning- of voltagegestuurde ionenkanalen
– ‘snelle’ ligandgestuurde ionenkanalen
– aan tyrosinekinase gekoppelde receptoren
– cytokine-receptoren.
Bij alle routes, behalve de eerste, wordt gebruik gemaakt van intracellulaire second messengers, waarvan de bekendste zijn: Na, K, Ca2, C1-, cyclisch AMP (cAMP), cyclisch GMP (cGMP), inositoltrifosfaat (1P3), diacylglycerol (DAG) en metabolieten van arachidonzuur (prostaglandinen, leukotriënen). Deze second messengers reguleren op hun beurt vaak de activiteit van andere routes van signaaltransductie, of van effectorenzymen, zoals proteïnekinasen (fosforylering van eiwitten) of fosfatasen (defosforylering van eiwitten). Het is in dit verband van belang op te merken dat de activiteit van vele enzymen bepaald wordt door de fosforyleringsgraad.
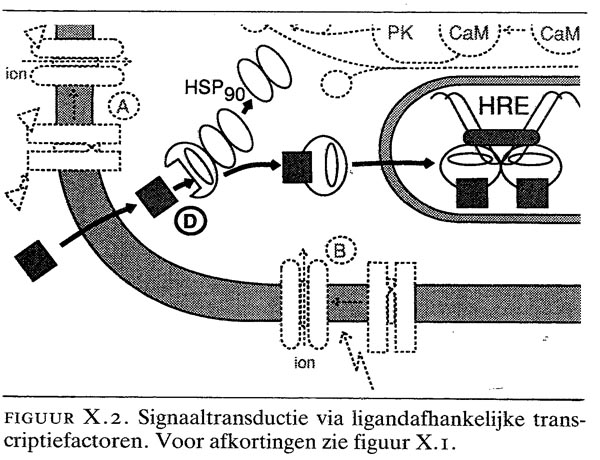
X.2. signaaltransductie via ligandafhankelijke transcriptiefactoren
Deze groep receptoren wordt ook wel de ‘steroïdreceptor-superfamilie’ genoemd. De liganden van deze groep receptoren zijn kleine, relatief apolaire verbindingen, zoals steroïde en thyroïdhormonen, retinoïnezuur (gevormd uit vitamine A) of vitamine D, die relatief gemakkelijk de celmembraan passeren (figuur X.2). In de cel binden de liganden aan receptoren, die daardoor geactiveerd worden. Door deze activatie wordt bijv. de cytoplasmatische glucocorticoïdreceptor ontkoppeld van zijn ‘chaperonne-molecuul’ en vervolgens naar de kern getransporteerd. Het werkingsmechanisme van hormoonantagonisten (bijv. tamoxifen, cyproteron, Ru 486) is dat deze wel aan hun respectieve receptoren binden, maar geen activatie induceren.
Receptoren bestaan uit een aantal functioneel autonome domeinen. Zo veroorzaakt één uiteinde van de steroïde receptoren de binding van de ligand, het middengedeelte de specifieke binding aan het regulatoire gebied van het doelgen (het ‘hormone response element’) en het andere uiteinde de daadwerkelijke modulering van de expressie van het doelgen. De structuur van het DNA-bindende middengedeelte vormt hèt karakteristieke kenmerk van de klasse van de steroïde receptoren.
Typerend voor ligandafhankelijke transcriptiefactoren is dat ze als dimeren op het DNA binden. Een hormone response element in het promotorgedeelte van een doelgen (zie hoofdstuk IV) bestaat dan ook altijd uit twee op elkaar lijkende stukken DNA. De binding van het hormoon-receptorcomplex als dimeer op het DNA is coöperatief: als het eerste receptormolecuul eenmaal gebonden is, wordt het tweede molecuul met een veel hogere affiniteit gebonden (tot 100 maal hoger). Door coöperatieve binding wordt het receptorcomplex stabieler.
X.3. signaaltransductie via aan g-eiwit gekoppelde receptoren
X.3.I . Receptoren
Kenmerkend voor deze superfamilie van receptoren in de plasmamembraan is dat de signaal transductie tussen het geactiveerde receptormolecuul en de intracellulaire effectoren verloopt via een guanosinetrifosfaat(GTP)-bindend regulatoir eiwit (figuur X.3). Aan de extracellulaire zijde van deze receptoren bevindt zich een nis waarin het ligand kan binden. Het intracellulaire deel van de receptor regelt de selectieve herkenning en activering van de verschillende typen G-eiwitten. Alleen een met ligand bezette receptor kan een G-eiwitcomplex binden en activeren. De intracellulaire effectoren van geactiveerde G-eiwitten zijn ionenkanalen, of enzymen die intracellulair second messengers genereren. Hoewel de aan G-eiwit gekoppelde receptoren één klasse vormen, kunnen ze een grote verscheidenheid aan liganden herkennen: polypeptiden, biogene aminen, eicosanoïden (metabolieten van arachidonzuur); ook licht- en reukreceptoren behoren tot deze superfamilie.
X.3.2. G-eiwitten
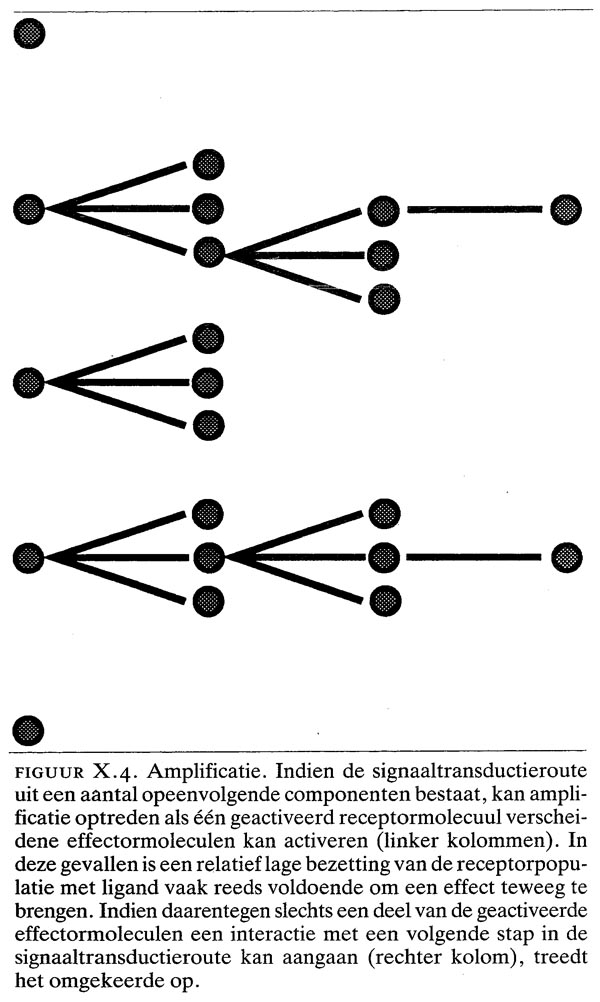
De G-eiwitten worden aan het binnenoppervlak van de plasmamembraan aangetroffen, waar ze functioneren als intermediair in de signaaloverdracht naar effector-eiwitten (zie figuur X.3b). Dit eiwit bestaat uit 3 subeenheden (?,? en ?). De ?-subeenheid, die het type G-eiwit bepaalt, bindt guaninenucleotiden, terwijl de hecht gebonden ??-dimeer aan de (plasma) membraan bindt. Onder niet-geactiveerde omstandigheden is de a-subeenheid aan de ??-dimeer gebonden en guanosine-difosfaat (GDP) aan de ?-subeenheid. Door interactie met een geactiveerde receptor wordt de ?-subeenheid geactiveerd: GDP wordt door GTP vervangen en het ?-GTP-complex dissocieert van de ??-dimeer. Eén geactiveerde receptor kan een aantal G-eiwitten activeren (amplificatie, figuur X.4). Een geactiveerd ?-GTP-complex kan op zijn beurt eveneens reageren met een aantal effector-eiwitten, zoals ionenkanalen, of enzymen die second messengers genereren. Het actieve ?-GTP-complex wordt geïnactiveerd door omzetting van GTP tot GDP. Het nu inactieve ?-GDP-complex associeert vervolgens opnieuw met het ??-complex (recycling).
Er bestaan vier functioneel verschillende typen ?-subeenheden. Een type stimuleert adenylaatcyclase, waardoor de intracellulaire concentratie van de second messenger cAMP toeneemt. Ditzelfde type ?-subeenheid kan ook Ca2-kanalen openen. Dit type G-eiwit wordt irreversibel geactiveerd door choleratoxine. Een tweede type a-subeenheid inactiveert daarentegen het adenylaatcyclase, sluit Ca2-kanalen, opent K-kanalen of activeert fosfolipase C. Na binding van pertussis-toxine kan dit type G-eiwit niet meer geactiveerd worden door een receptor. De andere typen a-subeenheden zijn pas onlangs gekarakteriseerd; deze regelen de stimulatie van de meer algemeen voorkomende, voor pertussis-toxine ongevoelige vorm van fosfolipase C.
X.3.3. Adenylaatcyclaseproteïnekinase A
Door binding van een geactiveerd ?-GTP-complex aan adenylaatcyclase wordt de omzetting van ATP in de second messenger cAMP geactiveerd (zie figuur X.3a). Cyclisch AMP activeert op zijn beurt het cytosolische enzym proteïnekinase A (PKA). PKA fosforyleert serine- of threonine-residuen in een eiwitketen. Een PKA-molecuul bestaat uit afzonderlijke regulatoire en katalytische subeenheden. Na binding van cAMP aan de regulatoire subeenheid verandert de conformatie hiervan, waardoor de katalytische subeenheid zich uit het complex kan losmaken en actief wordt.
X.3.4. Fosfolipasen
Een tweede, zeer belangrijk signaaltransductiepad is de activering van het membraan-gebonden enzym fosfolipase C (PLC) door ?-GTP-complexen (zie figuur X.3b). PLC splitst fosfolipiden tot diacylglycerol (DAG) en een polaire groep. Zo wordt fosfatidyl-inositoldifosfaat (PIP2) in DAG en inositoltrifosfaat (IP3) gesplitst. DAG en IP3 functioneren als second messengers voor respectievelijk de activering van proteïnekinase C (PKC) en de snelle mobilisering van intracellulair opgeslagen Ca2. Het ??-complex van G-eiwitten kan in een aantal gevallen fosfolipase A2 activeren, waardoor arachidonzuur wordt vrijgemaakt uit membraanlipiden. Uit arachidonzuur kunnen prostaglandinen en leukotriënen worden gevormd.
X.3.5. Proteinekinase C
De PKC-familie bestaat uit monomere enzymen met een gescheiden regulatoir en katalytisch domein (zie figuur X.3b). De binding van DAG aan het regulatoire domein veroorzaakt de activering van PKC. Doordat DAG snel gefosforyleerd wordt, is de activering van PKC door DAG kortdurend. Forbol-esters (krachtige co-carcinogenen) kunnen het effect van DAG nabootsen en PKC langdurig stimuleren. Evenals PKA fosforyleert PKC serine- en threonine-residuen in een eiwit.
Inactief PKC is niet membraan-gebonden. Het enzym kan pas geactiveerd worden na translokatie vanuit het cytosol naar de celmembraan. Fosfolipiden en Ca2 functioneren daarbij als essentiële cofactoren.
X.3.6. Ca2calmoduline-afhankelijke kinasen. Het door IP3 vrijgemaakte intracellulaire Ca2 wordt in het cytoplasma gebonden aan het eiwit calmoduline, dat daardoor van conformatie verandert (zie figuur X.3b). Geactiveerd calmoduline kan op zijn beurt onder andere Ca2calmoduline-afhankelijke proteïnekinasen activeren. Dit type kinasen, dat eveneens serine- en threonineresiduen fosforyleert, is in relatief hoge concentratie aanwezig in cellen met secretoire activiteit.
De cytosolische vrije Ca2-concentratie bedraagt slechts circa 100 nM, maar kan door IP3-stimulatie tot meer dan het 5-voudige stijgen. Door IP3 mobiliseerbaar Ca2 is intracellulair opgeslagen in een gespecialiseerd organel, het calciosoom. Dit organel is afkomstig is van het endoplasmatisch reticulum. Het sarcoplasmatisch reticulum in de spier is een gespecialiseerde vorm van het calciosoom. Opname van Ca2 in het organel vindt plaats via ATP-afhankelijke Ca2-pompen; Ca2 wordt opgeslagen als een complex met Ca2-bindende eiwitten om neerslag van calciumzouten in het organel te voorkomen. Mobilisatie en export van Ca2 naar het cytosol vinden plaats via binding van IP3 aan de IP3-receptor. Doordat IP3 snel geïnactiveerd wordt, is deze activering slechts tijdelijk. Het werkingsmechanisme van lithium berust mogelijk op een remming van de afbraak van IP3.
In een aantal celtypen reguleert een Ca2calmodulineafhankelijk proteïnekinase de vorming van het gas stikstofoxide (NO) uit arginine. NO is een zeer kort geleden ontdekte second messenger, die zich onderscheidt van andere second messengers door een intercellulaire werking: NO wordt in een ander celtype gesynthetiseerd dan waarin het zijn effect heeft. De intercellulaire werking komt tot stand via diffusie van het gas. In de aangrenzende cellen activeert NO het enzym guanylaatcyclase, waardoor cGMP accumuleert. Zo leidt de synthese van NO in het endotheel van de vaatwand tot relaxatie van de onderliggende gladde spieren. Het geneesmiddel nitroglycerine bootst deze werking van NO na.
X.3.7. ‘Langzame’ ligandgestuurde ionenkanalen
Het kenmerk van dit type receptoren, dat ook tot de aan G-eiwit gekoppelde receptoren behoort, is de relatief trage (0, 1-1 s) opening van ionenkanalen na binding van de ligand (zie figuur X.3c). Ionenkanalen kunnen rechtstreeks worden geactiveerd door de ?-subeenheid van G-eiwitten. Voorbeelden van dergelijke ionenkanalen zijn in het hart te vinden. Na ?-adrenerge stimulatie openen ?-GTP-complexen Ca2-kanalen (en activeren tegelijkertijd adenylaatcyclase). Stimulering van atriumcellen met acetylcholine activeert een type G-eiwitten, dat K-kanalen opent.
De licht- en reukreceptoren behoren tot een subtype van aan G-eiwit gekoppelde receptoren dat via de second messengers cGMP en cAMP de activiteit van kationenkanalen reguleert. Activering van de lichtreceptor rodopsine door licht leidt tot een verlaging van de intracellulaire cGMP-concentratie. Reukreceptoren zijn zeer verwant aan lichtreceptoren. Activering van de reukreceptor leidt tot intracellulaire accumulatie van cAMP. Daar sommige zoogdieren wel 10.000 verschillende geuren kunnen onderscheiden, is het niet verwonderlijk dat deze subfamilie van receptoren zeer uitgebreid is.
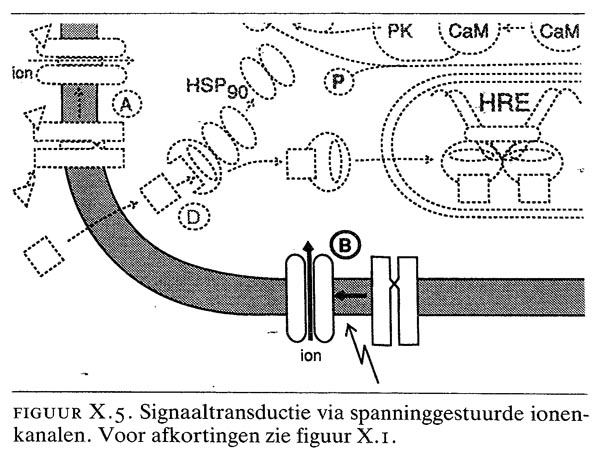
X.4. signaaltransductie via spanninggestuurde ionenkanalen
Spanning- of voltagegestuurde ionenkanalen openen onder invloed van een depolarisatie van de membraan (figuur X.5). Er zijn drie typen bekend: Na-, Ca2- en K-kanalen. Spanninggestuurde Ca2-kanalen zijn bijvoorbeeld betrokken bij de koppeling van excitatie en contractie van spieren en bij secretie door klieren. De activiteit van spanninggestuurde ionenkanalen wordt gereguleerd door kleine veranderingen in de membraanpotentiaal. De ‘spanningsensor’ wordt gevormd door positief geladen aminozuren in het ionenkanaal.
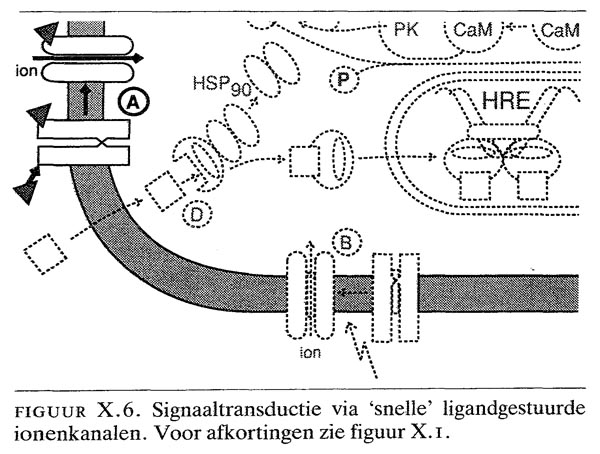
X.5. signaaltransductie via ‘snelle’ ligandgestuurde ionenkanalen
Kenmerkend voor dit type ligandgestuurde ionenkanalen is de zeer snelle opening van de kanalen (milliseconden) na binding van de ligand aan de receptor (figuur X.6). Zowel receptoren die kationen (o.a. Na) transporteren als receptoren die anionen (o.a. Cl-) transporteren, behoren tot deze familie. Bezetting van de ‘nicotine’-receptoren met acetylcholine opent bijv. Na-kanalen, waardoor een initiële depolarisatie optreedt. In de
motorische eindplaat van spieren worden hierdoor spanninggestuurde Na-kanalen geopend, waardoor de impuls zich voortplant en waardoor tevens Ca2 gemobiliseerd wordt ten behoeve van de spiercontractie. Nicotinereceptoren in de hersenen bepalen de uitwerking van geïnhaleerd nicotine op het gedrag. Bezetting van ?-aminoboterzuur- of glycine-receptoren leidt tot opening van Cl--kanalen, waardoor hyperpolarisatie van cellen optreedt. Deze receptoren, die vooral in het centrale zenuwstelsel vóórkomen en waarop onder andere benzodiazepinen en barbituraten aangrijpen, hebben dan ook vooral een negatief effect op de neuronale prikkeloverdracht. De ligandgestuurde ionenkanalen zijn ook gevoelig voor alcohol, waarbij de drempelwaarde slechts 1- 1,5‰ bedraagt.
X.6. signaaltransductie via aan tyrosinekinase gekoppelde receptoren
De receptor- en effectorfuncties van aan tyrosinekinase gekoppelde receptoren zijn verenigd in één molecuul (figuur X.7). Deze groep receptoren in de celmembraan is vooral betrokken bij proliferatie en differentiatie in dierlijke cellen. De liganden zijn meestal relatief kleine peptiden, zoals insuline en ‘platelet-derived growth factor’ (PDGF), terwijl de effectorfunctie bestaat uit fosforylering van tyrosine-residuen in eiwitketens. (De meeste proteïnekinasen fosforyleren serine- of threonineresiduen; slechts 0,1 van alle celeiwitten bevat fosfotyrosinen.) Binding van de ligand leidt tot dimerisatie van de receptor en daardoor tot activering van de tyrosinekinase-activiteit in het intracellulaire domein van de receptor. Als gevolg hiervan wordt dit intracellulaire deel ook zelf gefosforyleerd. Tot de gefosforyleerde eiwitten kunnen andere effector-eiwitten behoren, dat wil zeggen componenten van andere signaaltransductiepaden (bijv. fosfolipase C en diverse proteïnekinasen).
Een andere, niet-verwante familie van celmembraanreceptoren vormt die van de cytokine-receptoren. Binnen deze familie kunnen twee klassen worden onderscheiden. Tot de eerste klasse behoren de receptoren voor groeihormoon, prolactine, erytropoëtine, groeifactoren voor witte bloedcellen en de interleukinen, terwijl interferon-receptoren tot de tweede klasse behoren. Het mechanisme van deze signaaltransductie is nog grotendeels onbekend, hoewel goede aanwijzingen bestaan dat hiervoor associatie van de receptor met een tweede, intermediair signaaltransductiemolecuul nodig is. Sommige van deze receptoren (o.a. voor groeihormoon) kunnen ook zonder cytoplasmatisch en membraandomein worden gesynthetiseerd en uitgescheiden. Deze vormen circuleren als ‘carrier’-eiwit voor deze hormonen.
X.7. regulering van de signaaltransductie
Een receptor heeft twee functies: ligandherkenning en signaaltransductie. Een ligand moet niet met het substraat van een enzym vergeleken worden, maar met een allosteer werkende cofactor: door binding van de ligand verandert de conformatie van de receptor, die daardoor geactiveerd wordt en een signaal kan overdragen. De betrekkelijk geringe diversiteit van de mechanismen waarvan intracellulaire signaaltransductieroutes gebruik maken, contrasteert sterk met de relatief grote diversiteit van de signalen waarop een cel moet kunnen reageren. Eén signaal (ligand) wordt bovendien vaak herkend door verschillende receptorsubtypen. Zo zijn er van acetylcholine twee klassen van receptoren bekend (‘snelle’ ligandgestuurde ionenkanalen (nicotine-receptoren) en aan G-eiwit gekoppelde receptoren (‘muscarine’-receptoren)). Van de muscarine-receptoren zijn weer vijf subtypen bekend, die via verschillende signaaltransductiepaden werken (adenylaatcyclase en fosfolipase C). Omdat het effect van een signaal dus in de eerste plaats afhangt van het type receptor dat op of in een cel aanwezig is, staat het zoeken naar receptor-specifieke (ant)agonisten centraal in farmaceutisch onderzoek.
X. 7. 1. De betekenis van de affiniteit tussen receptor en ligand
Elk van de beschreven signaaltransductieroutes vormt een compromis tussen enerzijds het vermogen van de cel om lage concentraties te detecteren en anderzijds het vermogen snelle veranderingen in de concentratie van het extracellulaire signaal te registreren. De hoogste en de laagste dissociatieconstante (de ligandconcentratie waarbij de helft van het aantal beschikbare receptoren bezet is) van membraanreceptoren verschillen bijna een factor 106!
Een hoge dissociatieconstante (10-100 µM) betekent dat de receptor een geringe affiniteit voor de ligand heeft en daardoor zeer snel kan reageren op veranderingen in ligandconcentraties. Het voorbeeld van dit type receptoren is het ligandgestuurde ionenkanaal (zie figuur X.6), bijvoorbeeld de acetylcholine-receptor. De hoge ligandconcentratie die nodig is om zo'n receptor te activeren, kan alleen gedurende korte tijd in gespecialiseerde structuren (synapsen, motorische eindplaat) worden gerealiseerd. De gevoeligheid van dit type signaaltransductie wordt echter vergroot doordat receptoren vaak in zeer hoge concentratie in de postsynaptische membraan aanwezig zijn en slechts een deel van de receptoren geactiveerd hoeft te worden om het biologisch effect (bijv. depolarisatie, hyperpolarisatie) teweeg te brengen.
Een lage dissociatieconstante (0, 1-1 nM) betekent dat de receptor een zeer hoge affiniteit voor de ligand heeft. Hèt voorbeeld van dit type receptoren is de aan tyrosinekinase gekoppelde receptor (zie figuur X.7), bijvoorbeeld de insuline-receptor. Door de hoge affiniteit dissocieert het receptor-ligandcomplex relatief langzaam. Bij dit type receptoren is gevoeligheid voor lage concentraties van de ligand dan ook belangrijker dan het vermogen op snelle veranderingen in de ligandconcentratie te reageren.
Indien de signaaltransductieroute uit een aantal opeenvolgende componenten bestaat, ontstaat de mogelijkheid om een signaal te amplificeren. Amplificatie kan optreden als een relatief lage bezetting van de receptorpopulatie met ligand reeds voldoende is om een effect teweeg te brengen (zie figuur X.4). Amplificatie kan zinvol zijn als de efficiëntie van een volgende stap in een signaaltransductieroute gering is, bijvoorbeeld wanneer opeenvolgende stappen zich aan de binnenzijde van de celmembraan en in het cytosol afspelen (zie figuur X.4). Het voorbeeld van dit type signaaltransductie wordt gevormd door de systemen die gebruik maken van G-eiwitten als intermediair.
X. 7.2. Desensibilisering
Na een stimulering door hormonen of neurotransmitters vermindert de cellulaire gevoeligheid voor verdere stimulering door deze agentia gewoonlijk; men noemt dit desensibilisering. Desensibilisering treedt op doordat met ligand bezette receptoren tijdelijk van de celmembraan verwijderd worden door internalisering (via zgn. receptor-gemedieerde endocytose), of doordat receptoren ter plaatse geïnactiveerd worden door bijvoorbeeld fosforylering. Beide mechanismen zijn snel (tijdschaal: seconden tot minuten) en leiden niet tot een verandering van de totale hoeveelheid receptoreiwit in een cel. Dit is juist wel een kenmerk van ‘down regulation’, een mechanisme dat op veranderingen in synthese- en (of) afbraaksnelheid van de receptor en (of) een G-eiwit berust.
Er is een aantal specifieke receptorkinasen geïdentificeerd (zie figuur X.3a), die alleen met ligand bezette receptoren fosforyleren, en daardoor inactiveren. Interessant is dat deze receptorkinasen pas bij een relatief hoge bezettingsgraad van de receptor, zoals alleen in synapsen voorkomt, geactiveerd worden.
X. 7.3. Wisselwerking tussen signaaltransductiepaden
De specificiteit van het effect van een signaal kan worden vergroot door een onderlinge beïnvloeding van de intracellulaire signaaltransductiepaden. De beïnvloeding van de activiteit van een signaaltransductiepad door de effector van een ander signaaltransductiepad wordt ‘cross talk’ genoemd. Een voorbeeld hiervan is de activering van fosfolipase C door de tyrosinekinase-activiteit van de insuline-receptor (zie figuur X.7). Ook kunnen ‘cascades’ van een aantal effectorcomponenten van signaaltransductiepaden betrokken zijn bij het cellulaire antwoord op een signaal. Een voorbeeld hiervan vormen de proteïnekinasecascades, waarbij een geactiveerd kinase een ander type kinase activeert door dit te fosforyleren. Hierbij kan amplificatie optreden (zie figuur X.4). Veel van het hedendaagse celbiologische onderzoek is gericht op het ontrafelen van deze, vaak cel-specifieke, verknopingen van de signaaltransductiepaden.
Artikelinformatie
Citeer dit artikel als




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
(Geen onderwerp)
Amsterdam, februari 1992,
In het artikel van Lamers wordt een uitstekend overzicht gegeven van de verschillende aspecten die bij cellulaire signaaltransductie een rol spelen (1992;63-9). Naast de tyrosine-kinase-gekoppelde receptoren, onder andere voor diverse neurotrofe factoren,1 zijn er ook guanylaatcyclase- en proteïne-tyrosine-fosfatase-gekoppelde receptoren. Bovendien breidt de familie der cytokine- en neurokine-gekoppelde receptoren zich snel uit.2 Er zijn nu diverse liganden waarmee men in principe een aantal van de in het artikel genoemde receptoren in vivo kan onderzoeken met ‘single photon emission computerized tomograpy’ of ‘positron emission tomography’.3 Wellicht kan een goede samenwerking van preklinische en klinische wetenschappers leiden tot vraagstellingen voor in vivo-receptor-onderzoek bij de mens die van klinische belang zijn.
Thoenen H. The changing scene of neurotrophic factors. Trends Neurol Sci 1991; 14: 165-70.
Hall AK, Rao MS. Cytokines and neurokines: related ligands and related receptors. Trends Neurol Sci 1992; 15: 35-7.
Verhoeff NPLG. Pharmacological implications for neuroreceptor imaging. Eur J Nucl Med 1991; 18: 482-502.
Vaccinatie tegen influenza; aanmoedigen of aanzien?
s-Hertogenbosch, mei 1992,
In zijn brief werpt Salemink de vraag op of de waargenomen oversterfte tijdens de winterperiodes als gevolg van complicaties van influenza niet eigenlijk veroorzaakt zou kunnen zijn door een andere, gelijktijdig optredende oorzaak (1992;802). Uit het rapport Binnenjaarlijkse fluctuaties in de sterfte naar doodsoorzaak blijkt dat zowel influenza als koude (lage temperatuur) de fluctuaties in sterfte beïnvloedt.1 In anderhalve eeuw onderzoek is bij herhaling aangetoond dat influenza onafhankelijk van de buitentemperatuur leidt tot belangrijke oversterfte. Illustratief hiervoor zijn de epidemieën in (sub)tropische landen en de epidemieën van 1918-1919 en 1957,2 die in Nederland ook in de zomermaanden gewoed hebben. Voor alle duidelijkheid: de berekening van influenzaoversterfte, zoals in het proefschrift van Sprenger,3 geschiedt altijd na correctie voor seizoensinvloeden. De berekende oversterfte is dus netto, dat wil zeggen na aftrek van de jaarlijks in de winter terugkerende piek in sterfgevallen. Ook in het genoemde rapport werd nogmaals de noodzaak tot vaccinatie onder de risicogroepen aangetoond. Om de vaccinatiegraad van de risicogroepen te verhogen wordt door de Nederlandse Influenza Stichting dit jaar een voorlichtingscampagne gevoerd met als kern een Postbus 51-televisiespot.
Kunst AE, Looman CWN, Mackenbach JP. Binnenjaarlijkse fluctuaties in de sterfte naar doodsoorzaak. Rotterdam: Erasmus Universiteit, Instituut Maatschappelijke Gezondheidszorg, 1991.
Polak MF. Influenzasterfte in de herfst van 1957. [LITREF JAARGANG="1959" PAGINA="1098-109"]Ned Tijdschr Geneeskd 1959; 103: 1098-109.[/LITREF]
Sprenger MJW. The impact of influenza. Rotterdam, 1990. Proefschrift.
Vaccinatie tegen influenza; aanmoedigen of aanzien?
Amsterdam, juni 1992,
Collega Bouter et al. stellen mij gerust op het punt van de mogelijke verwarring met seizoensinvloed op de sterfte onder risicogroepen. Zij definiëren het begrip ‘oversterfte’ echter anders dan Meynaar et al. deden in het oorspronkelijk door mij becommentarieerde artikel (1992;168-72). Daar ging het om sterfte tijdens een influenza-epidemie aan andere oorzaken dan influenza en niet om oversterfte als gevolg van complicaties van influenza.
Vaccinatie van risicogroepen beperkt natuurlijk voor de betrokkenen het risico van influenza en de complicaties ervan, maar het verhindert niet het optreden van influenza-epidemieën en wij moeten dus maar hopen dat er een gunstige invloed van zal uitgaan op de steeds gelijktijdig optredende oversterfte aan andere oorzaken.