
artikel
Onderzoek gedurende de laatste eeuw heeft het mogelijk gemaakt tot in detail de ontwikkeling van de primaire en secundaire lichamelijke geslachtsverschillen te beschrijven. Onderzoek naar geslachtsverschillen van de hersenen is echter van recente datum bij het proefdier en nog maar net begonnen bij de mens. Pas in 1959 stelden Phoenix et al. door experimenteel onderzoek vast dat androgenen ook de differentiatie van de hersenen permanent beïnvloedden.1 Voordien waren door de eeuwen heen geslachtsverschillen in ziel en hersenen beschreven, maar meestal veeleer als afspiegeling van de maatschappelijke positie van de vrouw en haar reactie daarop, dan als resultaat van wetenschappelijk onderzoek. Zo zou volgens Aristoteles (384-322 v. Chr.) de mannelijke foetus bezield worden rond de 40e zwangerschapsdag en de vrouwelijke pas rond de 80e. Thomas van Aquino (1225-1274) nam deze opvatting zonder meer over. Hij was dan ook van mening dat de vrouw een “mas occasionatus” (een man die zijn bestemming niet heeft bereikt) zou zijn.
Ook de literatuur over geslachtsverschillen die in de vorm van de menselijke hersenen tot uiting komen, is een merkwaardige combinatie van “wetenschappelijke” observaties en culturele vooroordelen. Relatief kleine verschillen zijn herhaaldelijk gebruikt om de minderwaardigheid van de vrouw te bewijzen. Zo toonde Huschke in 1854 aan dat de voorhoofdskwab van de hersenen bij de man 1 groter is dan die van de vrouw, wat hem verleidde tot de verregaande uitspraak: “De vrouw is een homo parietalis en interparietalis, de man een homo frontalis, en de vorm van de hersenen van de vrouw is daarom ronder dan die van de man.” Hoewel Huschke toegaf dat het uitermate moeilijk was een geslachtsverschil in de hersenwindingen te vinden, stelde hij: er is echter geen twijfel mogelijk dat dit (verschil) bestaat.1 Mall gaf in een kritisch overzicht (1909) dan ook als zijn mening dat de verschillen gerapporteerd door Huschke voornamelijk berustten op de vooringenomenheid van de onderzoeker. Ook de gegevens van Retzius (1900) over geslachtsverschillen in hersengroeven en -windingen,1 werden al vroeg in twijfel getrokken, bijv. door Karplus (1905), die schreef: (...) Ik ben er momenteel niet van overtuigd dat uit de structuur van de windingen een minderwaardigheid van de vrouwelijke hersenen kan worden afgeleid.1 De waarneming van Meynert (1867) dat de man – in tegenstelling tot de vrouw – meer hersenweefsel vóór de centrale hersengroeve heeft dan erachter,1 werd bekritiseerd door Snell (1891) en Mall (1909), die verschillen in leeftijd en lichaamsgrootte in hun beschouwingen begonnen te betrekken.1 Mall (1909) concludeert dan ook: De bewering dat er specifieke (geslachts)verschillen in de hersenen bestaan, en dat de hersenen van de vrouw van een meer foetaal of aapachtig type zijn, zijn voornamelijk opinies zonder wetenschappelijke basis. Totdat wetenschappelijke onderzoekers specifieke verschillen aan kunnen tonen in gewicht en maat (. . .), zijn hun beweringen zonder wetenschappelijke waarde!1 Die tijd lijkt nu gekomen, want Malls bewering dat er geen variatie in de hersenbalk is in relatie tot (. . .) het geslacht, is onlangs ontzenuwd door De Lacoste-Utamsing en Holloway (1982), die dergelijke verschillen in vorm en afmeting hebben aangetoond.1 Het splenium was bij de vrouw meer bolvormig en het oppervlak was groter als de relatie tot het hersengewicht in aanmerking werd genomen. Dit grotere caudale gedeelte van het corpus callosum bij de vrouw komt overeen met de waarneming van McGlone (1981) en van Inglis en Lawson (1981) dat de twee hersenhelften bij de vrouw onderling meer functioneel verbonden zijn dan bij de man.1 Het idee dat de hersenen van de man een sterkere lateralisatie vertonen dan die van de vrouw is op zichzelf echter niet nieuw: Crichton-Browne stelde reeds in 1880 dat de neiging tot symmetrie van de twee hersenhelften bij de vrouw sterker is dan bij de man.1
Geslachtshormonen en hersenontwikkeling
De vele permanente geslachtsverschillen die de laatste decennia zijn aangetoond in de hersenen van dieren komen tijdens de periode van snelle hersenontwikkeling tot stand onder invloed van de geslachtshormonen die het zich ontwikkelende dier zelf aanmaakt. Eenvoudig voorgesteld: de aan- of afwezigheid van testosteron gedurende de fase van snelle hersenontwikkeling bepaalt de differentiatie van de hersenen in mannelijke of vrouwelijke richting. Bij de rat valt deze fase in de eerste weken na de geboorte, een periode die vergelijkbaar is met de hersenontwikkeling van de mens in de tweede helft van de zwangerschap en in de neonatale periode. In het mannelijke dier bepaalt testosteron door zijn effecten op celdeling, migratie, vorming van uitlopers en synapsen of de hersenen permanent in mannelijke richting differentiëren. Bij afwezigheid van testosteron in deze ontwikkelingfase differentiëren de hersenen in vrouwelijke richting. Door experimentele ingrepen in die fase kan men mannelijke hersenen in vrouwelijke richting laten differentiëren en omgekeerd. Men noemt dit proces de “organisatie” van de hersenen door geslachtshormonen. Een belangrijk punt is dat het masculiniserend effect van testosteron op de hersenen veelal tot stand komt nadat testosteron door de hersenen is gearomatiseerd tot oestrogenen. Dit betekent enerzijds dat experimenteel de effecten van testosteron ook met oestrogenen verkregen kunnen worden; anderzijds dat het geslachtsverschil in hersenontwikkeling dus blijkbaar meer een zaak is van de op dat moment aanwezige hoeveelheid geslachtshormoon dan van het soort hormoon.
Deze morfologische geslachtsverschillen in de hersenen zouden hun stempel drukken op het geslachtsverschil in gedrag. Het idee alleen al dat ook in dit opzicht de mens geen uitzondering zou vormen op de reeks onderzochte zoogdieren roept steeds weer bij velen sterke emoties op. Wellicht hebben deze niet zozeer betrekking op de mogelijk anatomische basis voor functionele geslachtsverschillen, zoals de gemiddelde superioriteit van de vrouw in bepaalde taalaspecten en de gemiddelde superioriteit van de man in ruimtelijke functies (bijv. McGlone 1980), of de geslachtsverschillen die zijn beschreven in het slaappatroon (Webb 1982) en de mate waarin de taalfuncties verdeeld zijn over de twee hersenhelften (McGlone 1980).1 Emoties komen vooral op wanneer de seksuele differentiatie van de hersenen centraal komt te staan in theorieën omtrent de ontwikkeling van onze hetero- of homoseksuele gerichtheid, of het ontstaan van een onbegrepen toestand als transseksualiteit. Bovendien wordt steeds weer de achterhaalde discussie gevoerd waarin het geslachtsverschil in absolute hersengrootte in direct verband wordt gebracht met een geslachtsverschil in intelligentie.
Ondanks de vele geslachtsverschillen van hersenfuncties die herhaaldelijk geconstateerd zijn en de vele dierexperimentele gegevens hierover, wordt de mogelijkheid van een “anatomische basis” in de hersenen van de mens voor zulke geslachtsverschillen nog steeds vaak fel verworpen. Het uitgangspunt en ideaal van de schrijver Couperus (1905), de “androgyne mens” – die evenveel mannelijke als vrouwelijke eigenschappen zou hebben1 – is gedurende de laatste tientallen jaren nieuw leven ingeblazen door de Women's Lib-beweging. Het bestaan van geslachtsverschillen wordt op vaak welhaast dogmatische wijze botweg ontkend voor zover het de hersenen betreft. Germaine Greer (1972) stelt bijv.: Hoewel geslachtshormonen de hersenen binnendringen (. . .) is er nooit een verband vastgesteld tussen dit fysiologische feit en geestelijke vermogens of gedrag.1
Geslachtsverschil in hersengewicht
Het geslachtsverschil in absoluut hersengewicht is inderdaad regelmatig gebruikt om de “minderwaardigheid” van de vrouw te “bewijzen”. Hoewel zowel feministen van de eerste lichting (suffragettes) als van de tweede (Women's Lib) volkomen terecht afkeurend hebben gewezen op dit misbruik, hebben bekende vrouwen uit beide periodes dezelfde fouten gemaakt bij het “bewijzen” van de “superioriteit” van de vrouw. Een vroeg voorbeeld kan gevonden worden in het handboek van Maria Montessori (1913).1 Zij schrijft: Er heerst zoals u weet een wijdverbreid geloof dat bevestigd wordt uit naam van de wetenschap: dat de vrouw biologisch – met andere woorden totaal – minderwaardig is, dat het volume van haar hersenen door de natuur veroordeeld is tot een minderwaardigheid waartegen niets bestand is (. . .). Dat het hersenvolume moet worden bezien in relatie tot de lichaamsgrootte is een bekend principe. Zo vergelijkt Manouvrier de hersenen met het lichaamsgewicht (. . .). Hierna volgt een berekening die niet in detail te achterhalen noch te begrijpen valt, waaruit Maria Montessori concludeert: Het hersenvolume van de vrouw is dus superieur aan dat van de man!, (wat niet juist is – zie fig. 1). Zij vervolgt: Dit is een antropologische superioriteit die verder ook blijkt uit de meer perfecte vorm van de schedel, in zoverre dat de vrouw een absoluut recht voorhoofd heeft en dus geen spoor van bogen boven de oogkas (een kenmerk van superioriteit van de soort). Zo bestaat er dus een tegenstelling tussen antropologische en sociale omstandigheden: de vrouw die uit antropologisch standpunt bezien de schedel heeft van een welhaast superieur ras, wordt nog steeds zonder meer gedegradeerd tot een sociaal minderwaardige positie, waaruit zij moeilijk te verheffen valt.1
De toegenomen belangstelling van het hersenonderzoek voor geslachtsverschillen valt samen met de tweede feministische golf, wat heeft geleid tot een versterkte kritiek vanuit de Women's Lib-beweging op dit type onderzoek. Er werd op gewezen dat, vooral in de huidige economische recessie, aspecten van dit onderzoek gebruikt zouden kunnen worden om de dominante positie van de man te handhaven als “een biologisch bepaalde onontkoombaarheid”. Deze vrees zou kunnen verklaren (zonder haar te rechtvaardigen!) waarom bijv. Germaine Greer (1972) een soortgelijke redeneertrant gebruikt om een “vrouwelijke superioriteit” aan te tonen:1 Er werd gedacht dat de relatief lichtere vrouwelijke hersenen een argument zouden zijn voor een geringere geestelijke capaciteit, hoewel is aangetoond dat vrouwen in verhouding tot hun lichaamsgewicht zwaardere hersenen hebben (dit is niet juist – zie fig. 1). De schrijfster vervolgt: Hoe dan ook, zodra het hersengewicht in het nadeel van de man begon uit te vallen, werd het niet meer van belang geacht. Als de voorhoofdskwab gezien moet worden als de zetel van de intelligentie, dan moet er ook op worden gewezen dat dit gebied meer ontwikkeld is bij de vrouw. (Een referentie hiervoor wordt door Greer niet gegeven, hoewel Huschke (1854) een tegenovergesteld verschil vond van slechts 1,1 en Mall (1909) in deze kwab geen geslachtsverschil vond.)1
Hersengrootte en intelligentie werden overigens niet uitsluitend met elkaar in verband gebracht als het over geslachtsverschillen gaat. Röse (1905), die een aantal Duitse professoren en soldaten onderzocht, kwam tot de slotsom: “De professoren hebben aanzienlijk grotere schedels dan de officieren. De gewone hoogleraren hebben de grootste schedel, op de voet gevolgd door de overige universitaire docenten: buitengewoon hoogleren (. . .).1 Bayerthal (1911) vervolgde met de opmerking: Men kan met een minimale schedelomtrek van 52-53 cm tenminste nog hoogleraar in de chirurgie of verloskunde worden, maar met een schedelomtrek van minder dan 52 cm kan men geen intellectuele prestaties van enige betekenis verwachten, terwijl onder de 50,5 cm geen normale intelligentie kan worden verwacht. In dit verband maakte hij de opmerking: We hoeven niet te vragen naar de schedelomtrek van vrouwelijke genieën – die zijn er niet.1
Geslachtsverschil in relatieve hersengrootte; allometrie
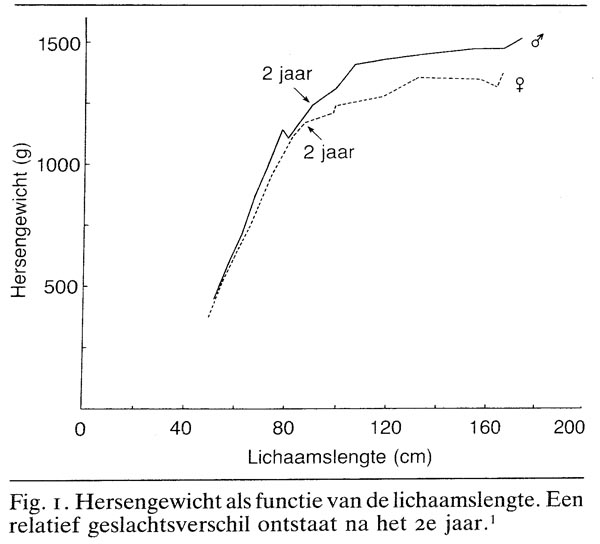
Sedert de tweede helft van de negentiende eeuw is bekend dat een geslachtsverschil in absoluut hersengewicht reeds aanwezig is bij de geboorte (Bischoff 1880; Pfister 1897); de recente literatuur is hiermee in overeenstemming, hoewel de gevonden waarden verschillen. De schedelomtrek laat een consistent verschil zien van zo'n 2-3.1 Echter, niet alleen de hersenen van pasgeboren jongetjes zijn zwaarder dan die van meisjes, ook hun lichaamsgewicht is hoger. Deze relaties zijn echter meestal niet te leggen op grond van de simpele verhouding: hersengewicht lichaamsgewicht. Veeleer worden ze het beste beschreven door een niet-lineaire, exponentiële allometrische vergelijking. Het uitwerken van dit soort relaties wordt allometrie genoemd, een terrein dat gedefinieerd is als de studie van de grootte en haar consequenties. Uit een grafische weergave van de relatie tussen hersengewicht en lichaamslengte (een maat die niet gevoelig is voor de hoeveelheid vetweefsel, in tegenstelling tot het lichaamsgewicht) blijkt dat het geslachtsverschil bij de geboorte inderdaad wegvalt (zie fig. 1). Pasgeboren meisjes en jongetjes hebben dus, indien ze dezelfde lichaamslengte hebben, ook gemiddeld hetzelfde hersengewicht. Een relatief geslachtsverschil ontstaat echter rond het tweede jaar en blijft gedurende het gehele leven bestaan.1 De mogelijk functionele betekenis van dit geslachtsverschil is thans niet bekend.
Geslachtsverschil in microscopische bouw van de hersenen
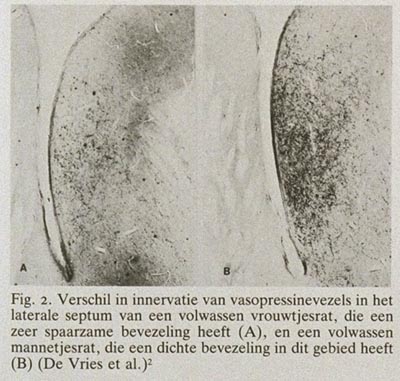
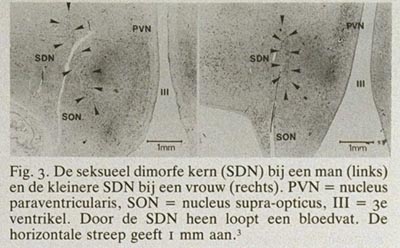
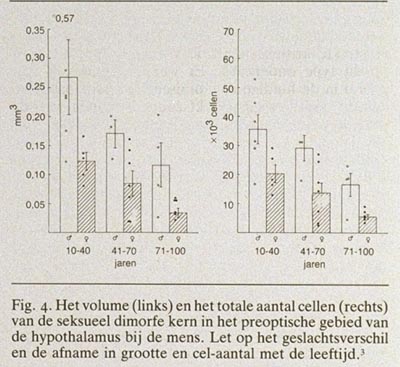
Met het blote oog valt vooral de overeenkomst in bouw op tussen mannelijke en vrouwelijke hersenen, maar dat wil niet zeggen dat er geen microscopische of biochemische verschillen zouden bestaan. Uit recent immunocytochemisch onderzoek wordt duidelijk dat door het gehele dierenrijk heen hersenstructuren, die er met de conventionele kleurtechnieken hetzelfde uitzien, uit cellen en vezels zijn opgebouwd die per geslacht zeer verschillend kunnen zijn. Als voorbeeld kan dienen de kleuring van een boodschapper in de hersenen, het neuropeptide vasopressine. De vasopressine-innervatie van het laterale septum bij de rat is bij mannetjes zeer uitgesproken en bij vrouwtjes bijna niet aanwezig (fig. 2). Dergelijke immunocytochemische technieken voor de tientallen chemische boodschappers in de hersenen worden momenteel toepasbaar gemaakt voor menselijk hersenweefsel en zullen de komende tijd zeker ook gegevens opleveren over mogelijke chemische geslachtsverschillen. Microscopisch zijn bij zoogdieren geslachtsverschillen gemeten in alle denkbare morfologische aspecten van het zenuwstelsel (celaantal, dendrietbomen, synapsverdelingen, neurotransmitters, etc.). Het duidelijkste geslachtsverschil in de structuur van zoogdierhersenen is beschreven door Gorski et al. (1978) en betreft een celgroep in het pre-optisch gebied.1 Deze celgroep vertoont bij de rat zo'n duidelijk cyto-architectonisch geslachtsverschil dat het met het blote oog zichtbaar is na een conventionele Nissl-kleuring. Zeer onlangs hebben wij een analogon van deze “seksueel dimorfe nucleus” (SDN) in het pre-optisch gebied van de hypothalamus bij de mens gevonden.3 Morfometrische analyse liet zien dat de SDN bij de man 2,5 maal zo groot is als bij de vrouw en 2,2 maal zo veel cellen bevat (fig. 3 en 4). Er wordt momenteel nog gewerkt aan de allometrische relatie tussen het volume van de SDN en het totale hersengewicht bij de mens. De eerste resultaten lijken er op te wijzen dat een kleine toename in het hersengewicht een sterke, exponentiële toename van het volume van de SDN tot gevolg heeft. Dit zou kunnen verklaren waarom dit kerngebied bij mannen, gezien hun wat grotere absolute hersengewicht, zo aanzienlijk groter is dan bij vrouwen. Hoewel de functie van de SDN nog niet bekend is, ligt deze kern in een gebied dat in het dierexperiment essentieel is voor de regulatie van gonadotrope hormonen en seksueel gedrag.
Geslachtshormonen en hersenontwikkeling bij de mens
Onderzoek met moderne technieken naar geslachtsverschillen in de bouw van de menselijke hersenen is nog maar van zulke recente datum dat er thans geen uitspraken te doen zijn over de relatie tussen zulke geslachtsverschillen en gedrag, hetero- of homoseksuele gerichtheid of transseksualiteit. Wel is uit neuropsychologisch onderzoek reeds duidelijk dat geslachtshormonen ook bij de mens tijdens de ontwikkeling in principe permanente gedragsveranderingen teweeg kunnen brengen. Zo'n mechanisme is voor het eerst geopperd naar aanleiding van vermannelijking van aspecten van het gedrag van meisjes met het adrenogenitale syndroom. Het blijkt echter ook uit effecten van geslachtshormonen die door artsen toegediend zijn, bijv. in geval van een dreigende miskraam. Niet dat geslachtshormonen een miskraam zouden voorkomen, maar wij artsen geven nu eenmaal graag iets. Geslachtshormonen als diëthylstilbestrol (DES), die toegediend worden aan de zwangere moeder, passeren echter naar het kind, dat hierdoor later een geringe kans (ca. 1:10.000) op een vaginacarcinoom heeft en een grote kans op functionele ontwikkelingsstoornissen van de tractus genitalis die tot voortplantingsproblemen leiden (in ca. 25). Bovendien beïnvloeden geslachtshormonen waarschijnlijk de hersenontwikkeling van het kind. Diverse veranderingen in seksueel dimorf gedrag (d.w.z. gedrag waarin mannen en vrouwen van elkaar verschillen) zijn namelijk waargenomen bij kinderen van wie de moeder tijdens de zwangerschap oestrogenen, progestativa of combinaties hiervan kreeg toegediend.45 Prenataal toegediende progestativa met androgene werking verhogen later de scores in agressietests bij meisjes en jongens (Reinisch 1981). Andere progestativa zouden het gedrag van jongens juist feminiseren en de libido verminderen, terwijl aan DES blootgestelde jongens zich een meer “conventionele” mannelijke jeugd herinnerden.6 Tijdens de zwangerschap aan DES blootgestelde meisjes vertoonden later een meer mannelijke lateralisatie in een verbale test7 en een toename van “stereotiep” vrouwelijk gedrag.8 Recent Amerikaans onderzoek liet zien dat DES-dochters een verhoogde kans op biseksualiteit en homoseksualiteit hebben.9 DES is niet alleen door artsen verstrekt. Het was in Nederland ook per postorder verkrijgbaar als seks-stimulerend middel. Het zal wel nooit te achterhalen zijn hoeveel zwangere vrouwen dit middel om deze reden hebben gebruikt.
Hoewel dit soort waarnemingen belangrijke informatie kan geven over de seksuele differentiatie van de menselijke hersenen en het gedrag, is het te hopen dat deze onbedoelde experimenten snel zullen stoppen. Recent dierexperimenteel onderzoek maakt echter duidelijk dat de geslachtelijke differentiatie tijdens de ontwikkeling van de hersenen niet alleen beïnvloed wordt door geslachtshormonen, maar ook door stoffen die de neurotransmitters dopamine of acetylcholine beïnvloeden, door barbituraten en andere geneesmiddelen, door alcohol of door stress van het moederdier.10 Geneesmiddelen worden door de industrie in het algemeen niet onderzocht op hun invloed op de seksuele differentiatie,11 zodat de seksuele differentiatie van de hersenen van het kind ook door vele tot nu toe nog niet genoemde chemische stoffen ongewild beïnvloed kan worden. Hierdoor, en door de recente mogelijkheid structuren die geslachtelijke differentiatie vertonen in de menselijke hersenen immunocytochemisch te kleuren en te meten, zal er in de komende decennia dus waarschijnlijk voldoende materiaal beschikbaar komen voor dit nieuwe onderzoeksterrein naar hersenstructuren en de factoren die onze geslachtelijke ontwikkeling in de ruimste
zin van het woord bepalen.
Literatuur
Swaab DF, Hofman MA. Sexual differentiation of the humanbrain. A historical perspective. In: Vries GJ de, Bruin JPC de, Uylings HBM,Corner MA, eds. Sex differences in the brain. Prog Brain Res 1984; 61:361-74.
Vries GJ de, Buijs RM, Swaab DF. Ontogeny of thevasopressinergic neurons of the suprachiasmatic nucleus and theirextrahypothalamic projections in the rat brain – presence of a sexdifference in the lateral septum. Brain Res 1981; 218: 67-78.
Swaab DF, Fliers E. A sexually dimorphic nucleus in thehuman brain. Science 1985; 228: 1112-5.
Ehrhardt AA, Meyer-Bahlburg HFL. Effects of prenatal sexhormones on gender-related behavior. Science 1981; 211: 1312-8.
Yalom ID, Green R, Fisk N. Prenatal exposure to femalehormones. Effect on psychological development in boys. Arch Gen Psychiatry1973; 28: 554-61.
Kester P, Green R, Finch SJ, Williams K. Prenatal femalehormone administration and psychosexual development in human males.Psychoneuroendocrinology 1980; 5: 269-85.
Hines M, Shipley C. Prenatal exposure todiethylstilbestrol (DES) and the development of sexually dimorphic cognitiveabilities and cerebral lateralization. Developmental Psychology 1984; 20:81-94.
Ehrhardt AA, Meyer-Bahlburg HFL, Feldman JF, Ince SE.Sex-dimorphic behavior in childhood subsequent to prenatal exposure toexogenous progestogens and estrogens. Arch Sex Behav 1984; 13:457-77.
Ehrhardt AA, Meyer-Bahlburg HFL, Rosen LR, et al. Sexualorientation after prenatal exposure to exogenous estrogen. Arch Sex Behav1985; 14: 57-75.
Swaab DF, Mirmiran M. Mechanisms underlying thebehavioral teratogenic effects of chemicals on the developing brain. In:Flehmig I, ed. Proceedings of the 2nd European symposium on Developmentalneurology. Stuttgart: Gustav Fischer Verlag, 1985.
Kenyon P, Malinek V. Screening drugs for effects onsexual differentiation. Neurobehav Toxicol Teratol 1984; 6:1-2.
Artikelinformatie
Citeer dit artikel als



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Reacties